2. Definition of study object
2.1 Synapsida
2.2 Therapsida
2.3 Dinocephalia
2.4 Titanophoneus
2.5 Titanophoneus potens Efremov
Краткая систематика объекта:
Надкласс Tetrapoda
Класс †Синапсиды (
Synapsida)
Подкласс †
Therapsida Инфракласс †Эотерапсиды (Eotherapsida)
Отряд †Диноцефалы (Dinocephalia)
Инфраотряд †Антеозавры (Anteosauria)
Надсемейство †Антеозавроидеи (Anteosauroidea)
Семейство †Антеозавровые (Anteosauridae)
Род †Титанофонеи (Titanophoneus)
Вид †T. potens
Вид †T. adamanteus
Вид †T. rugosus
Общий вид системы дан по M. F. Ivakhnenko. 2003. Eotherapsids from the East European Placket (Late Permian). Paleontological Journal 37(Suppl. 4):S339-S465 [R. Butler/R. Butler]
А теперь об этих и других новостях подробнее.
2.1 Synapsida
читать дальше
Группа Synapsida была выделена относительно недавно, в конце XX века. Поскольку в дальнейшей работе будут отмечены некоторые общие особенности данной группы, стоит дать подробную характеристику.
До конца ХХ века среди надкласса Tetrapoda \\Четвероногие// выделяли четыре класса: Амфибии (вместе с надклассом Рыбы сотавляют группу Анамнии), Рептилии, Птицы и Млекопитающие (вместе эти три класса составляют группу Амниоты). В источниках того времени Synapsids are group of mammal-like reptiles lived during Permian period which is extinct now . \\синапсиды -- группа зверообразных рептилий, живших в пермский период и ныне вымершая.// Синапсид тогда рассмтривали как подкласс в классе Reptilia. Надо вхуяить в текст ссылку на конкретный источник
После выхода работ немецкого энтомолога Willi Hennig оформилось такое направление систематики как кладистика. Кладистика есть способ построения естественной системы, который предлагает строгое соответствие между уровнем родства организмов и положением групп в иерарии системы. При этом, в отличии от эволюционной таксономии, кладистика требует строгой монофилии. Клада -- основная единица в кладистике -- это группа животных, ведущих происхождение от одного предка (это и есть требование монофилии) и включающая всех потомков этого предка.
При дальнейшем распространении кладистики пришлось серьёзно пересмотреть систематику тетрапод. Reptiles in the traditional sense are a paraphyletic group \\ Рептилии в традиционном их значении -- группа парафилитическая. // То есть, включает в себя существ, происходящих от общего предка (клювоголовых, крокодилов, чешуйчатых), но не всех, птицы рассматривались как отдельный класс. Однако птицы отделились от динозавров много позже того времени, как динозавры разделились на семейства, стило быть, являются живущей ныне группой динозавров. Традиционная филогенетическая систематика допускала это. Всем было похуй. Но с распространением кладистики потребовалось либо разделить класс рептилий на на несколько (и повысить ранг существующих отрядов рептилий до уровня подкласса), либо ввести дополнительную иерархическую единицу, объединяющую птиц и рептилий в одну кладу и противопосталяющую их сестринской группе. Либо крестик снимите, либо трусы наденьте!
В настоящее время класс Рептилий устарел, рептилии -- это не систематическая группа, а эволюционный уровень, града. Ну-ка, поднимите руки те, у кого в школьных учебниках написано про класс рептилий? Вот так и живём, образование совсем не поспевает за наукой. Для разделения эволюционных уровней были предложены клады Амниот (it's a clade of tetrapod vertebrates who have embryotic membranes formed as eggs)//(такая клада наземных чтвероногих позвоночных, у которых есть набор зародышевых оболочек, формирующих яйца)// и анамний (это парафилитическая группа, в которую входят позвоночные животные, не имеющие зародышевых оболочек).
Амниоты, появившиеся в конце каменноугольного периода, примерно 310 миллионов лет назад (1) относительно быстро разделились на две ветви: Завропсиды Sauropsida (к ним как раз относятся Анапсиды (ныне вымершая группа) и Диапсиды (к диапсидам относятся все нынеживущие классы рептилий и птицы) и Sinapsidamorpha (к этой группе относятся млекопитающие и не-маммальные ранние синапсиды). Хотя, по правилам кладистики, млекопитающие также являются синапсидами, они имеют выраженные апоморфные признаки по сравнению с не-маммальными синапсидами. К примеру, для разделения млекопитающих и прочих синапсид используется такой диагностический признак млекопитающих: «сочленение нижней челюсти с черепом, при котором сочленовная головка зубной кости входит в гленоид (суставную впадину) на чешуйчатой кости»
Здесь и в дальнейшем термин "синапсид" буду использовать в значении "ранний синапсидоморф", разделяя дочерний класс млекопитающих и предковую группу "не-маммальных синапсид". Примерно так же как название "динозавр" используется для сокращения, вместо длинного термина "нептичий динозавр" по отношению к группам, побочным, по отношению к птичей линии. Группа Sinapsida для удобства "мерджа" с традиционой систематикой будет упоминаться в ранге класса. (Хотя современная кладистическая систематика тяготеет к безранговости, введение традиционных таксономических единиц бывает удобным).
Я вот тут подумал мимоходом, если бороться за высокую культуру и быт, то надо было бы иметь кладу Anapsida и какую-нить Apsidia. И в Апсид включить диапсид и синапсид, а то всякие анапсиды хронологически раньше отделились от общего ствола. Но на эту тему пускай сами кладисты головы и ломают, я и так шибко долго экскурс в систематику пишу.
Главням признаком, определяющим синапсид является наличие одного височного отверстия в черепе. У анапсид вообще нет скуловых дуг и височных отверстий, их череп почти монолитный, а у диапсид развиваются две скуловые дуги и два височных отверстия: верхнее и нижнее. Верхнее височное окно хорошо заметно на черепах диапсид в dorsal view //вид со спины//.
Рис.1. Представители клад Sauropsida и Diapsida (фотографии автора)
Височное отверстие черепа у синапсид образовано костями: скуловой, заглазничной, чешуйчатой, квадратно-скуловой. У Титанофона строение височного отверстия типичное для ранних синапсид.
Другая важная черта синапсид это раннее воникновение в этой эволюционной ветви гетеродонтии. Уже на уровне пеликозавров у представителей рода Dimetrodon есть разделение зубов по функциям, что и нашло отражение в названии этого животного: "диметродон" означает "две меры (два типа) зубов". У представителей подкласса Therapsida уже есть дифференциация на три типа зубов, характерных также и для млекопитающих: резцы, клыки и заклыковые зубы. Заклыковые зубы у большинства представителей терапсид мелкие, примитивной конической формы. В некоторых группах (цинодонты) заклыковые зубы имеют достаточно сложную форму. В дальнейшем наметился эволюционный тренд на стабилизацию формы и количества зубов в челюсти и формирование окклюзии (координированного смыкания зубов верхней и нижней челюсти, наподобии ножниц). Окклюзия резцов возникала в разных группах довольно рано, окклюзия коренных зубов, также возникавшая независимо, способствовала возникновению нового типа обработки пищи во рту: жевания. Считается, что первыми животными, способными к жеванию (т.е. боковым движениям челюсти, способствующим перетиранию пищи) стали представители Anamodontia.
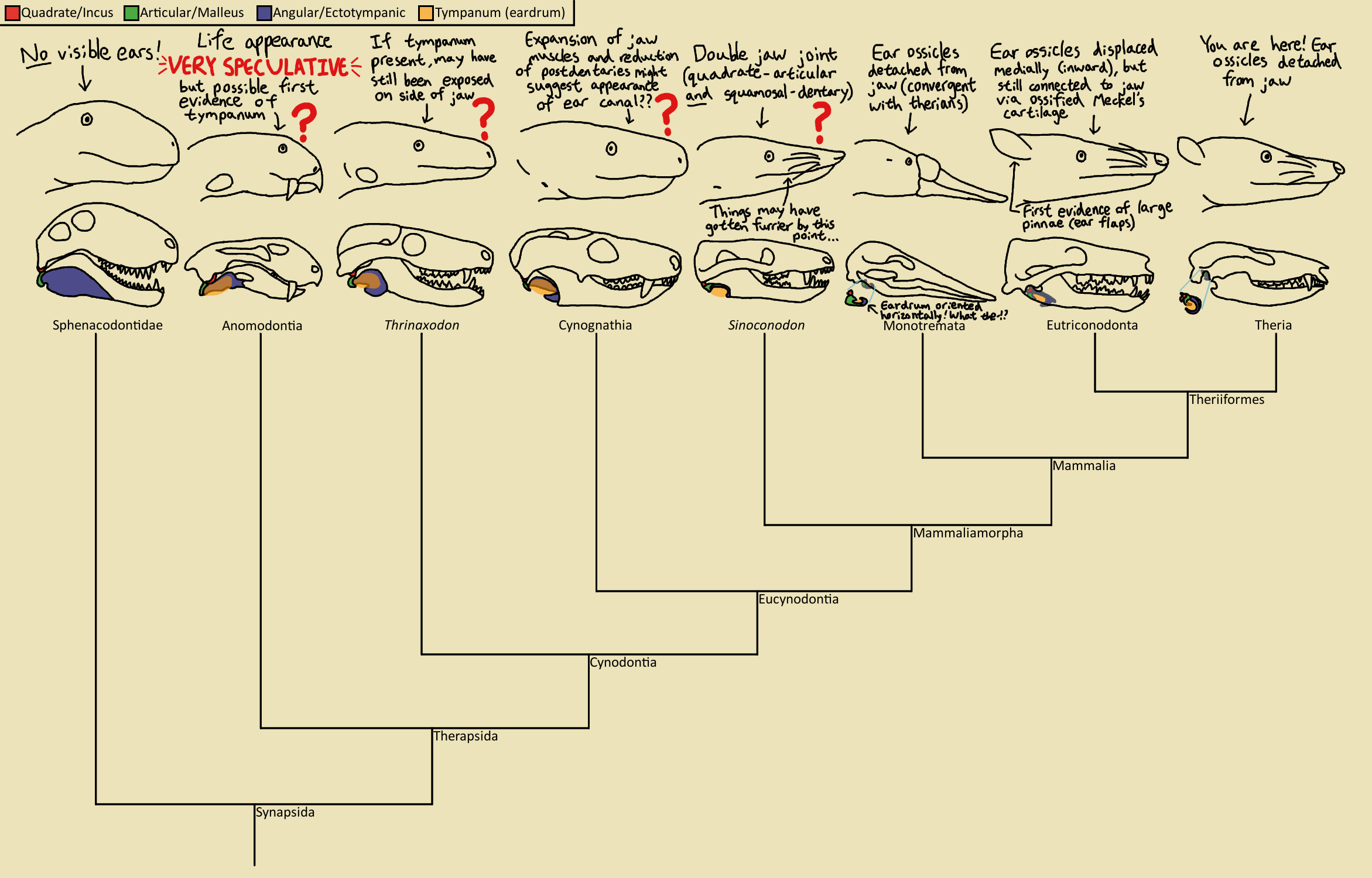
В ходе эволюции нижняя челюсть синапсид постепенно теряла кости, расположенные сзади от зубной кости dentale, которая в свою очередь неуклонно увеличивалась, разрастаясь назад. Далее она пришла в соприкосновение с чешуйчатой костью черепной коробки. В результате возникло новое сочленение нижней челюсти с черепом. Это сочленение, а также преобразование среднего уха как раз и являются рубежом, отделяющим "немаммальных синапсид" от синапсид-млекопитающих.
большая картинка
Череп синапсид в целом имеет относительно монолитную структуру, по сравнению с диапсидным, имеющим облегчённую ажурную структуру. Это позволило сформировать прочную черепную коробку и, в эволюционной перспективе, стало преадаптацией к дальнейшему развитию головного мозга, впрочем, реализованному гораздо позже, на уровне плацентарных млекопитающих.
Уже на уровне пеликозавров заметны "эксперименты" синапсид по ускорению обмена веществ. У диметродона на спине был внушительный кожный парус, позволявший активнее получать солнечное тепло на манер солнечной батареи. Для более поздних синапсид была показана возможность наличия диафрагмы, а, следовательно, и активного дыхания. В монографии Татаринова указывается трактовка крупных кровеносных сосудах на некоторых черепах как кровеносных сосудов, питающих вибриссы. А для поздних тероцефалов вязниковского комплекса было в явном виде показано наличие шерстяных покровах в работах польских и российских палеонтологов (3).
Различные фауны синапсид встречаются на всех континентах, бывших в конце пермского периода объединённых в единый суперконтинент -- Пангею. Именно синапсиды первыми в истории Земли создали в пермском периоде богатую и разнообразную фауну позвоночных суши. Среди пермских синапсид были крупные сильные хищники размером с тигра, с огромными кинжаловидными клыками; были охотники на мелкую добычу, аналоги куньих и псовых; существовали и растительнояды, большие тяжеловесные животные, а уже на уровне млекопитиющих им удалось освоить жизненную форму мелких фитофагов.
Класс Synapsida подразделяется на группы Pelycosauria и Therapsida, которые также являются сборными и отображают скорее градами, чем кладами. В традиционных системах им присваивается ранг подкласса (Ивахненко) или отряда.
Литературка:
1. Benton M. J., Donoghue P. C. J., Asher R. J., Friedman M., Near T. J., Vinther J. Constraints on the Timescale of Animal Evolutionary History // Palaeontologia Electronica, 2015, 18.1.1FC. — P. 1—106.
2. Benton M. J. Vertebrate Paleontology. 3rd ed. — Oxford: Blackwell Science Ltd., 2005. — 750 p. — ISBN 0-632-05637-1. — P. 393—399.
3. Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia Doi: 10.1111/let.12156
4.Palaeoneurological clues to the evolution of defining mammalian soft tissue traits опубликована в журнале Scientific Reports
Doi: 10.1038/srep25604